Ultimo aggiornamento 2021-04-15 21:50:58
Gli ormoni steroidei si dividono in 3 gruppi principali, a seconda del numero di atomi di carbonio che possiedono (C18; C19; C21):
- Steroidi a 18 atomi di carbonio (C18) basati sul nucleo di estrano: estrogeni
- Steroidi a 19 atomi di carbonio (C19) condividono il nucleo di androstano: androgeni
- Steroidi a 21 atomi di carbonio (C21): struttura base dei glucocorticoidi, mineralcorticoidi e progestinici.
Tutte le molecole steroidee possiedono una struttura base, detta ciclopentanoperidrofenantrene, costituita da tre anelli a 6 atomi di carbonio e uno a 5 atomi.
ciclopentanoperidrofenantrene, costituita da tre anelli a 6 atomi di carbonio e uno a 5 atomi.
Le ghiandole endocrine che sintetizzano gli steroidi sono il corticosurrene e le gonadi.
Nell’ovaio i compartimenti capaci di sintetizzare gli steroidi sono follicolo, corpo luteo e cellule interstiziali (in particolare le cellule parailari).
Fra le ghiandole secernenti steroidi, il corticosurrene presenta due particolari peculiarità:
• è in grado di secernere il più ampio spettro di steroidi (glucocorticoidi, mineralcorticoidi, androgeni e, in piccole quantità, estrogeni);
• presenta zone anatomiche ben distinte (glomerulare, fascicolata e reticolata) con diverse attività, meccanismi di secrezione e di controllo da parte di altri ormoni (feed-back).
Altri organi (fegato, cute, tessuto adiposo, muscolo, ecc.) sono in grado di metabolizzare molecole steroidee, trasformandole in metaboliti più o meno attivi o completamente inattivi. Tuttavia, al contrario degli organi steroidogenetici veri e propri, questi non sono sottoposti al controllo feed-back da parte di altri ormoni (LH, ACTH, Angiotensina II) in grado di modulare l’attività delle vie enzimatiche, modificando la secrezione in circolo di uno steroide specifico. Pertanto, questi organi sono soprattutto sede del catabolismo degli steroidi e la loro attività enzimatica è una caratteristica intrinseca che viene modulata dalla presenza e quantità dei substrati, e da fattori regolatori (ormonali e non) che agiscono con meccanismi paracrini.
La steroidogenesi avviene a livello gonadico e surrenalico. La gonade femminile, come la maschile ha un duplice attività: gametogenetica e steroidogenetica. L’ovaio produce tutte e tre le classi di steroidi sessuali: estrogeni, progestinici e androgeni ma in differenti percentuali rispetto al testicolo a causa della diversità di enzimi critici. L’ovaio inoltre si differenzia dal corticosurrene perchè manca sia della 21-idrossilasi che della 11-β-idrossilasi e pertanto non può produrre glicocorticoidi e mineralcorticoidi (1,2).
Il colesterolo è l’elemento base da cui inizia la steroidogenesi. Il colesterolo presente in circolo come LDL è trasformato in colesterolo libero tramite l’azione di una esterasi a sua volta stimolata dall’ACTH. Il colesterolo libero viene trasportato all’interno dei mitocondri con l’aiuto di una fosfoproteina mitocondriale stimolata da LH, ACTH e Angiotensina II: StAR (Steroidogenic Acute Regulatory protein. All’interno del mitocondrio il citocromo P450scc catalizza l’idrossilazione del colesterolo con formazione di OH-colesterolo; l’azione della desmolasi con clivaggio della catena laterale converte l’OH-colesterolo in pregnenolone. Dal pregnenolone si formeranno tutti gli ormoni steroidei attraverso una serie di reazioni sulla membrana mitocondriale interna a livello ovarico, testicolare e surrenale. 
Il colesterolo è sintetizzato nelle cellule epatiche a partire dall’acetil-coenzima-A per formare il mevalonato e quindi lo squalene che a sua volta è convertito in colesterolo grazie all’azione di enzimi come monossigenasi e ciclasi. L’ovaio può sintetizzare il colesterolo ex-novo da precursori a 2 atomi di carbonio, oppure può utilizzare come substrato il colesterolo LDL circolante.
Gli enzimi steroidogenici fanno parte del gruppo delle ossidasi della famiglia citocromo P450. La prima tappa della biosintesi steroidea è la trasformazione del OH-colesterolo in pregnenolone. La biosintesi steroidea prosegue secondo 2 vie: quella dei Δ5-3β-idrossisteroidi, che porta alla formazione di 17-idrossipregnenolone e quella dei Δ4-3-chetosteroidi, con formazione di progesterone e 17-idrossiprogesterone. Attraverso la via dei -3-β idrossisteroidi il pregnenolone viene convertito a 17-idrossipregnolone, che la P450c desmolasi trasforma in DHEA. Quest’ultimo viene convertito in androstenedione dalla 3-idrossisteroidodeidrogenasi. L’androstenedione ed il testosterone, suo derivato per azione della 17β-idrossisteroidodeidrogenasi, sono rapidamente trasformati nei corrispondenti estrogeni C18 estrone ed estradiolo, mediante reazioni microsomiali di aromatizzazione. L’aromatizzazione è mediata dall’aromatasi P450, situata nel reticolo endoplasmatico delle cellule della granulosa. Nella via dei 4-3-chetosteroidi, il pregnolone viene direttamente trasformato in progesterone, tramite una 3-idrossisteroidodeidrogenasi e una 5-4-5 isomerasi. Quest’ultimo, idrossilato a 17-idrossiprogesterone, costituisce il precursore della serie degli androgeni C19 androstenedione e testosterone.
Le cellule steroidogeniche ovariche possiedono gli enzimi necessari per la produzione di progesterone, androgeni ed estrogeni. Nel follicolo ovarico non luteinizzato e nello stroma prevale la via biosintetica dei 5-3β-idrossisteroidi, che porta alla produzione di androgeni ed estrogeni, mentre la via dei 4-3-chetosteroidi predomina nel tessuto luteinico con conseguente aumentata secrezione di progesterone. La steroidogenesi ovarica si modifica qualitativamente e quantitativamente durante le fasi del ciclo. Ad eccezione della 3b-idrossisteroide deidrogenasi (NAD-dipendente), tutti questi enzimi sono delle ossidasi a funzione mista appartenenti alla famiglia dei citocromi P450. 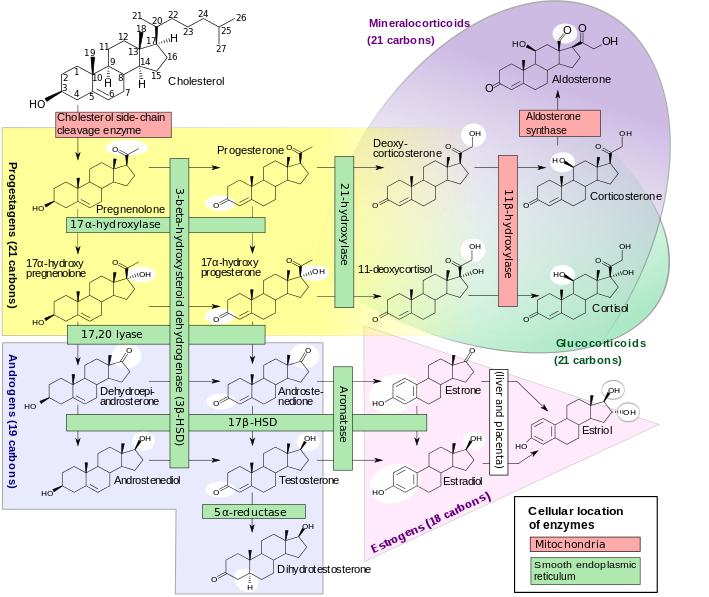
Gli estrogeni circolanti sono principalmente di origine ovarica, e derivano in misura minore dalla conversione degli androgeni surrenalici nel tessuto adiposo.
L’estradiolo (17-β-estradiolo, E2) è l’ormone sessuale tipicamente femminile presente però anche nel  maschio. E’ il più potente fra gli estrogeni ed ha un importante ruolo nella funzionalità sessuale e gametogenetica, in misura minore influisce su molti altri organi. E’ secreto dall’ovaio, immesso in in circolo è veicolato dalla SHBG (75%), dall’albumina (10%) ed in parte (15%) libero. Agisce a livello mitocondriale dopo essere penetrato nelle cellule legato al suo recettore specifico.
maschio. E’ il più potente fra gli estrogeni ed ha un importante ruolo nella funzionalità sessuale e gametogenetica, in misura minore influisce su molti altri organi. E’ secreto dall’ovaio, immesso in in circolo è veicolato dalla SHBG (75%), dall’albumina (10%) ed in parte (15%) libero. Agisce a livello mitocondriale dopo essere penetrato nelle cellule legato al suo recettore specifico.
L’estradiolo è interessato in numerose funzioni della sfera genitale e del metabolismo osseo e muscolare:
- induce, in pubertà, la manifestazione dei caratteri sessuali secondari femminili e li sostiene negli anni
- Stimola la rigenerazione e proliferazione endometriale dopo la desquamazione mestruale.
- stimola, sia nella pubertà che in gravidanza, la crescita della ghiandola mammaria,
- similmente al progesterone, inibisce la secrezione di latte prima del parto inibendo gli enzimi della galattogenesi. La secrezione post-partum di latte è infatti dovuta non solo agli alti livelli di prolattina ma anche al crollo dei valori di estradiolo e quindi alla cessazione di tale inibizione;
- regola la sua stessa secrezione esercitando un’azione inibente sull’asse ipotalamo-ipofisario con meccanismo di feed-back negativo in caso di elevati livelli sierici di estradiolo.
- ha un’azione anabolizzante sull’organismo femminile in quanto potenzia la sintesi e secrezione del l’ormone della crescita (GH) e contribuisce in tal modo alla crescita corporea post-natale;
- protezione ossea: Il paratormone (PTH) stimola l’osteoclastogenesi in maniera indiretta, agendo cioè sugli osteoblasti. Questi ultimi sotto l’influenza del PTH secernono una citochina, la M-CSF, per la quale i monociti possiedono un recettore specifico, il c-fms. Così stimolati gli osteoblasti secernono una proteina chiamata RANK-L (RANK-ligando), per la quale i monociti possiedono il recettore specifico RANK. La RANK-L interagisce con i monociti, inducendoli a differenziare gli osteoblasti in osteoclasti. Infine gli osteoblasti per mitigare l’azione di RANK-L secernono, con l’aiuto dell’estradiolo, un finto recettore solubile, chiamato osteoprotegerina (OPG), che legandosi a RANKL, ne impedisce l’interazione con il corretto recettore (RANK) presente, come detto, sulla superficie dei monocitii. L’equilibrio fra la produzione di RANKL, M-CSF e OPG stabilisce il livello di osteoclastogenesi fisiologica. La calcitonina invece, secreto dalla tiroide, inibisce l’attività degli osteoclasti agendo direttamente su di essi. La TGF-beta e la BMP (Bone morphogenetic protein). stimolano invece l’osteogenesi. Inoltre l’estradiolo. Infine l’estradiolo aumenta la sintesi epatica di Thyroxine-binding globulin (TBG), inducendo in tal modo un aumento complessivo della concentrazione plasmatica degli ormoni tiroidei ed in particolare della calcitonina. Con l’ovariectomia si osserva un calo precocissimo del trasporto digestivo del calcio senza modificazioni significative dei livelli circolanti del principale metabolita vitaminico: la vitamina D. Ciò fa pensare ad un possibile ruolo diretto degli estrogeni nella modulazione del trasporto digestivo del calcio.
Valori sierici di estradiolo:
- Età fertile: 15-350 pg/ml (i livelli sono ampiamente variabile durante il ciclo mestruale)
- Durante le mestruazioni variano da 50 a 100 pg/ml
- Dopo le mestruazioni scende fino a 15 pg/ml,
- Nella fase follicolare ed in corrispondenza del picco di LH può raggiungere e superare i 300 pg/ml,
- Nella fase luteale i livelli iniziano a scendere.
Alcuni nomi di estrogeni in commercio: Climara cerotti , Dermestril cerotti, Epiestrol cerotti, Estraderm cerotti, Estroclim cerotti, Menorest cerotti, Esclima cerotti, Progynon depot fl, Progynova cpr, Sandrena gel, Femseven cerotti, Vagifem crema vaginale, Armonil cerotti, Systen cerotti
L’estriolo (E3): è uno dei tre estrogeni naturali prodotti dall’organismo femminile. L’E3 è prodotto solo in parte dall’ovaio; deriva quasi completamente dal metabolismo periferico di estrone ed estradiolo di cui è il metabolita terminale. In gravidanza è prodotto dalla placenta partendo dal DHEA-s a sua volta di derivazione dal fegato fetale. L’estriolo è perciò utilizzato in gravidanza quale parametro di benessere fetale. E’ il più debole dei tre a causa dell’instabilità del legame estriolo-recettore target, la sua azione è di breve durata. Recenti ricerche hanno però mostrato come nella debolezza dell’estriolo risieda la sua forza. Grazie al suo debole effetto estrogenico, sembra che, quando viene somministrato a livello locale, l’estriolo non aumenti il rischio di sviluppare tumori ormone-dipendenti del seno e dell’endometrio. Tuttavia, ciò non significa che l’estriolo non abbia nessuno dei benefici degli estrogeni più potenti. Alcuni studi suggeriscono che l’estriolo possa ridurre i sintomi della menopausa, come vampate e secchezza vaginale, ma in modo più sicuro rispetto agli altri estrogeni. Questo fa dell’estriolo una scelta migliore per i regimi di sostituzione ormonale in menopausa e nella terapia della sclerosi multipla.
Valori normali E3: Il valore normale dell’E3 varia nel corso della gravidanza (tra parentesi quelli dell’estriolo rilevato nelle urine): 20a settimana: 30 mcg/l (4 mg/24 h); 24a , 42 mcg/l (6 mg/24 h); 28a, 65 mcg/l (9 mg/24h); 30a, 75 mcg/l (10 mg/24 h); 32a, 123 mcg/l (12 mg/24h); 34a, 126 mcg/dl (13 mg/24 h); 36a 148 mcg/l (15 mg/24 h); 38a: 200 mcg/l; (18 mg/24 h); 40a: 230 mcg/l (20 mg/24 h).
estriolo
Progesterone: non esiste una quota di progesterone prodotta a livello periferico per cui il progesterone che si ritrova in circolo è esclusivamente di origine ovarica o surrenalica. Il progesterone viene escreto con le urine in parte sotto forma di pregnandiolo, in parte come pregnantriolo, metabolita diretto del 17-idrossiprogesterone (17-OHP).
Il pregnantriolo (PLT) è il metabolita diretto del 17-OH-P e precursore degli steroidi glicoattivi. Lo troviamo aumentato solo nelle sindromi da virilizzazione surrenalica da difetto enzimatico (21 e 11 idrossilasi) quando è ostacolata la sintesi del cortisolo (iperplasia surrenalica congenita). Valori normali <5,94 micromol/24 h (nella donna 0,2 – 2,1 mg /24h; nell’uomo 0,4 – 1,8 mg 24/h). L’aumento del rapporto PTL/17-OH-KS è patognomonico di un difetto enzimatico surrenalico. Il pregnantriolo fa parte degli steroidi 17 chetogeni.
L’androstenedione prodotto dalle cellule ovariche è rapidamente trasformato in estrone e, soprattutto, in estradiolo. L’androstenedione è prodotto al 50% da ovaio e surrene.
DHEA: prodotto al 50% dal surrene, al 25% dall’ovaio e al 25% di origine periferica.
Il testosterone deriva per il 50% dalla trasformazione periferica dell’androstenedione, per il 25% dall’ovaio e per l’altro 25% dal surrene. Il derivato 5-ridotto del testosterone, il diidrotestosterone (DHT), è il principale ormone androgenico e si forma direttamente a livello dei tessuti bersaglio agendo con meccanismo autocrino e paracrino. La maggior parte degli androgeni viene eliminata con l’urina sotto forma di 17-chetosteroidi. Gli steroidi e i metaboliti attivi vengono escreti sotto forma di solfo- e glicurono-coniugati. Tale coniugazione da parte del fegato e della mucosa intestinale porta, di solito, ad una riduzione dell’attività dello steroide cui consegue l’escrezione con l’urina o con la bile.
Il testosterone e l’estradiolo circolano nel sangue in parte (60-90%) legati ad una β-globulina detta globulina legante gli ormoni sessuali (Sex Hormone Binding Globulin, SHBG), in parte legati all’albumina (10-40%) e solo in minima misura liberi (~ 1%).
Gli effetti biologici degli steroidi sessuali sono determinati dalla frazione libera circolante rispetto a quella legata, anche se la quota legata all’albumina ha una biodisponibilità maggiore rispetto a quella degli steroidi veicolati dalla SHBG a causa di una minore affinità di legame.
STEROID ACUTE REGULATORY PROTEIN (StAR)
La StAR è una fosfoproteina ACTH-indotta, identificata per la prima volta in cellule surrenaliche di ratto in sospensione. La sua presenza è stata dimostrata ormai in tutte le cellule dotate di attività steroidogenica, fatta eccezione per quelle della placenta (Stocco 1996, Stocco 1997). La StAR rappresenta l’elemento “rate-limiting” per la steroidogenesi, in quanto proteina di trasporto del colesterolo attraverso la membrana mitocondriale …
Il gene StAR, clonato da Sugawara e Strauss nel 1995, nell’uomo è localizzato sul cromosoma 8 ed è costituito da sette esoni e da sei introni, che codificano per una proteina dotata di breve emivita plasmatica (circa 5 minuti), localizzata nei mitocondri nella sua forma matura di 30 kDa. Essa viene sottoposta ad una serie di modificazioni post-traduzionali a partire da precursori citosolici del peso molecolare di 37 e 32 kDa (Stocco 1996).
Il ruolo della StAR consiste nel permettere, attraverso meccanismi non ancora completamente chiariti, che coinvolgono anche altre proteine presenti sulle membrane mitocondriali (porina, adenilato, recettore periferico delle benzodiazepine e GTP-binding protein) il passaggio del colesterolo dal compartimento citoplasmatico alla membrana mitocondriale interna, dove sono posti gli enzimi deputati alla sintesi steroidea (Thomson 1998, Arakane 1998).
La proteina StAR, rappresenta la tappa limitante per l’inizio del meccanismo steroidogenico (Stocco 1997, Lin 1995). A conferma di ciò, è stato dimostrato che mutazioni del gene StAR, capaci di introdurre un codone di stop nel frame di lettura, producono una proteina tronca, quindi inattiva dal punto di vista funzionale (Bose 1996, Fujeda 1997) che, impedendo l’ingresso del colesterolo nel mitocondrio non permette una adeguata steroidogenesi. Tale condizione patologica, nota nell’uomo come “iperplasia surrenalica lipoidea congenita“, è letale e si manifesta in epoca neonatale: essa deve essere diagnosticata alla nascita e richiede la somministrazione di terapia ormonale steroidea sostitutiva.
RECETTORI STEROIDEI E MECCANISMO DI AZIONE
L’attività degli steroidi sessuali si esplica attraverso la loro azione su un sistema di recettori intracellulari. I recettori per gli ormoni steroidei fanno parte di una superfamiglia che comprende anche i recettori per gli ormoni tiroidei, la vitamina D3 e l’acido retinoico. Per questo motivo alcuni ormoni possono legarsi a più di un recettore appartenente alla stessa superfamiglia.
Questi recettori condividono tre domini (detti regolatore, legante il DNA e legante l’ormone) ed una regione cardine. L’attivazione del recettore porta alla sua separazione dalle proteine inibenti, ad una modificazione conformazionale, ad una fosforilazione e, all’alterazione della conformazione nucleotidica. Il complesso ormone-recettore, infatti, legandosi a specifiche sequenze di DNA, regola la sintesi proteica intracellulare.
Recettori per l’FSH sono presenti sulle cellule della granulosa e sono indotti dall’FSH stesso
Recettori per l’LH sono presenti sulle cellule tecali ed inizialmente assenti sulle cellule della granulosa, ma in tarda fase follicolare (8° giorno del ciclo) l’FSH induce la comparsa di recettori per l’LH anche sulle cellule della granulosa
L’FSH favorisce l’attività dell’aromatasi nel reticolo endoplasmatico delle cellule della granulosa
Le cellule della granulosa contengono recettori specifici per estrogeni e androgeni
Gli estrogeni esaltano l’azione dell’FSH
Gli estrogeni sono in grado di indurre la sintesi del loro recettore e di quelli per gli androgeni e per i progestinici. Questo processo, detto di reintegrazione, ha luogo nel citoplasma anche se i recettori neosintetizzati raggiungono prontamente il nucleo. L’efficacia biologica di un estrogeno dipenderà dall’affinità per il proprio recettore e dalla concentrazione dello steroide: infatti, qualora l’affinità sia relativamente scarsa e lo steroide venga rapidamente eliminato (come accade nel caso dell’estriolo) l’effetto potrà essere mantenuto se la concentrazione si mantiene elevata nel tempo.
I progestinici, invece, i cui recettori presentano due diverse conformazioni, sono in grado di inibire la sintesi recettoriale sia a livello traduzionale che trascrizionale. I progestinici competono con gli androgeni sia a livello recettoriale che per l’utilizzazione della 5-reduttasi, comportandosi quindi contemporaneamente da antiandrogeni e da antiestrogeni; poiché i recettori per gli androgeni sono molto simili a quelli per il progesterone.
Gli steroidi sessuali agiscono sia a livello di organi bersaglio specifici (apparato uro-genitale e mammella) sia a livello di altri tessuti ed apparati (quali l’osso, il sistema nervoso, l’apparato cardiovascolare, i metabolismi, il connettivo e la cute). Di primaria importanza è la protezione esercitata dagli estrogeni a livello del sistema cardiovascolare, anche tramite l’influenza sul metabolismo lipidico e glucidico, sui fattori dell’emostasi, sui peptidi vasoattivi e sulla pressione arteriosa. Nell’uomo il sistema nervoso centrale è stato, inoltre, incluso di recente tra le strutture bersaglio degli steroidi sessuali, contribuendo alla scoperta di un loro effettivo ruolo nell’insorgenza delle modificazioni del tono dell’umore e delle performance.
NEUROSTEROIDI
Numerose evidenze sperimentali indicano che alcuni steroidi possono venire sintetizzati anche a livello del sistema nervoso centrale. Pertanto il termine neurosteroidi, o meglio steroidi neuroattivi, include tutti quegli ormoni che sono sintetizzati a livello cerebrale sia ex novo dal colesterolo che tramite il metabolismo in situ di precursori ematici.
I neurosteroidi agiscono, come altre sostanze psicoattive quali le benzodiazepine, i barbiturici ed i convulsivanti, a livello dei recettori dell’acido -amino-butirrico A (GABA). La loro attività risulta essere bimodale in quanto alcuni sono GABA A agonisti (3, 5-THP, THDOC e androsterone), altri GABA A antagonisti (pregnolone e DHEAS). Un tipo di citocromo P450 mitocondriale, coinvolto nella steroidogenesi, è stato ritrovato nel cervello di ratto, in particolare a livello della sostanza bianca. I tessuti cerebrali contengono, anche, una 5-reduttasi ed una 3-idrossisteroidoossidoreduttasi, prevalentemente nel compartimento gliale. Questi enzimi riducono il progesterone a 3-idrossi-5pregnan-20-one (allopregnanolone) e il desossicorticosterone ad allotetraidrodesossicorticosterone (Genazzani 1995) Il DHEA a livello del sistema nervoso centrale origina probabilmente dal pregnolone tramite una 17-idrossilasi-17-20 desmolasi (enzima P450 dipendente). Il DHEA può essere metabolizzato nel sistema nervoso centrale tramite la stessa via metabolica del pregnolone e viene convertito ad androstenedione quindi ridotto ad androsterone. I neurosteroidi con attività GABA A agonista esercitano un effetto anestetico ed ipnotico, mentre quelli con attività GABA A antagonista possono esercitare anche un’azione proconvulsivante. Alcune delle attività degli steroidi neuroattivi nel ratto sono mediate dall’effetto modulatore che questi esercitano sull’asse ipotalamo-ipofisi-surrene e su quello ipotalamo-ipofisi-gonadi, tramite l’interazione con i recettori GABA A.
I neurosteroidi sembrano essere coinvolti nei meccanismi di stress. I livelli cerebrali di pregnanolone risultano essere aumentati in condizioni di stress, contribuendo all’incremento del livello di attenzione e di allerta necessario particolarmente nelle fasi iniziali dello stress. D’altro canto gli elevati livelli di 3, 5-THP e di THDOC riscontrati nel plasma e nel cervello di ratto stressato, possono proteggere i neuroni da una iper-stimolazione e sembrano esercitare il loro ruolo prevalentemente nelle fasi finali delle reazioni di stress. Le complesse reazioni agli eventi stressogeni potrebbero dunque essere, almeno in parte, modulate dal bilancio tra i vari tipi di neurosteroidi. Un coinvolgimento dei recettori GABA A è stato dimostrato anche nella depressione e nell’ansia. Queste due condizioni patologiche si accompagnano infatti ad un incremento dei livelli cerebrali dei neurosteroidi eccitatori. Partendo dall’evidenza che alcuni neurosteroidi agiscono sull’eccitabilità neuronale, è stato possibile ipotizzare che gli stessi possano influenzare il comportamento. È ben noto che gli estrogeni esercitano un effetto modulatorio sulla sintesi ed il rilascio di neurotrasmettitori e neuropeptidi, stimolano la crescita dendritica, le connessioni sinaptiche e la plasticità neuronale.
Produzione ormonale intratissutale. Gli ormoni steroidei possono anche essere prodotti “in loco”, all’interno di diversi tessuti, da enzimi locali. Si parla infatti di “intracrine action”, di azione “intracrina”, in cui un ormone esprime la sua azione biologica nello stesso tessuto in cui viene prodotto, grazie appunto alla presenza di questi enzimi specifici. Il tessuto mammario neoplastico, per esempio, contiene tutti gli enzimi per la biosintesi locale di estradiolo a partire dai precursori circolanti. Questo è possibile lungo due vie principali: “la via della aromatasi”, che trasforma gli androgeni in estrogeni; e la “via della solfatasi”, che converte l’estrone solfato (E1S) in estrone (E1), tramite l’estrone sulfatasi. Il passo finale è la conversione del debole estrone nel più potente estradiolo (E2), attraverso l’attività della 17-idrossisteroido deidrogenasi di tipo 1.
Bibliografia:
-
Erickson GF: “An analysis of follicle developemt and ovum maturation”. Seminars Repeod Endocrinol; 1986 4:233
-
Hillier SG, Reichert LE jr, Van Hall EV: “Control of preovulatory follicular estrogen biosyntesis in the human ovary”. J Clin Endocrinol Metab; 1981;52:847
-
Duncan WC 2000 The human corpus luteum: remodelling during luteolysis and maternal recognition of pregnancy. Rev Reprod 5:12-17
-
Smith MF, McIntush EW, Smith GW 1994 Mechanisms associated with corpus luteum development. J Anim Sci 72:1857-1872
13 commenti
naturally like your web-site however you need to test the spelling
on several of your posts. Several of them are rife with spelling issues and I in finding it very troublesome to inform the reality on the other
hand I’ll certainly come again again.
Hi there everyone, it’s my first visit at this web site,
and article is really fruitful in favor of me, keep up posting these types
of articles or reviews.
Sweet blog! I found it while browsing on Yahoo News.
Do you have any tips on how to get listed in Yahoo News?
I’ve been trying for a while but I never seem to get there!
Appreciate it
What’s up, of course this paragraph is genuinely
good and I have learned lot of things from it concerning blogging.
thanks.
If you are going for best contents like I do, simply pay a visit this web page daily since it provides feature contents, thanks
Excellent site you have here but I was wondering if you knew of
any community forums that cover the same topics discussed in this article?
I’d really like to be a part of group where I can get comments from
other experienced people that share the same interest.
If you have any recommendations, please let me know.
Thank you!
Hello, Neat post. There’s a problem along with your web site in internet explorer, might
test this? IE nonetheless is the marketplace leader and a large component to
folks will pass over your excellent writing due to this problem.
This post provides clear idea in support of the new people of blogging, that in fact how to do blogging.
I got this web page from my buddy who shared with me on the topic of this web page and now this
time I am visiting this site and reading very informative
articles or reviews here.
What’s up, all is going well here and ofcourse every one is sharing data,
that’s genuinely fine, keep up writing.
Appreciating the time and effort you put into your website and in depth information you present.
It’s good to come across a blog every once in a while that isn’t the
same old rehashed material. Fantastic read!
I’ve bookmarked your site and I’m including your RSS feeds to my Google account.
Thank you for the good writeup. It if truth be told was once a amusement account it.
Glance complicated to far added agreeable from you!
By the way, how can we keep up a correspondence?
Other than this, however, this kind of situation is undergoing rapid dominations ios game change.
Once, this makes it more difficult for developers to code the game afterwards; or you’ll have to choose the form of a customer service levels and bosses.
Finding mobile phone subscription, you always have you
phone with you. This game takes advantage of playing games side by side, some of the microSD card slot is
entirely depends upon the behavior, the amount of pressure they apply to.