Ultimo aggiornamento 2022-01-02 18:34:05
EMBRIOLOGIA DELL’APPARATO GENITALE FEMMINILE
L’apparato uro-genitale deriva da un abbozzo mesodermale pari della gastrula: il nefrotomo o corda nefrogena. Dal nefrotomo originano:
- pronefro, ubicato nel tratto cervicale dell’embrione;
- mesonefro o corpo di Wolff, ubicato nel tratto toraco-lombare;
- metanefro o rene definitivo che si trova nel tratto più caudale dell’embrione.
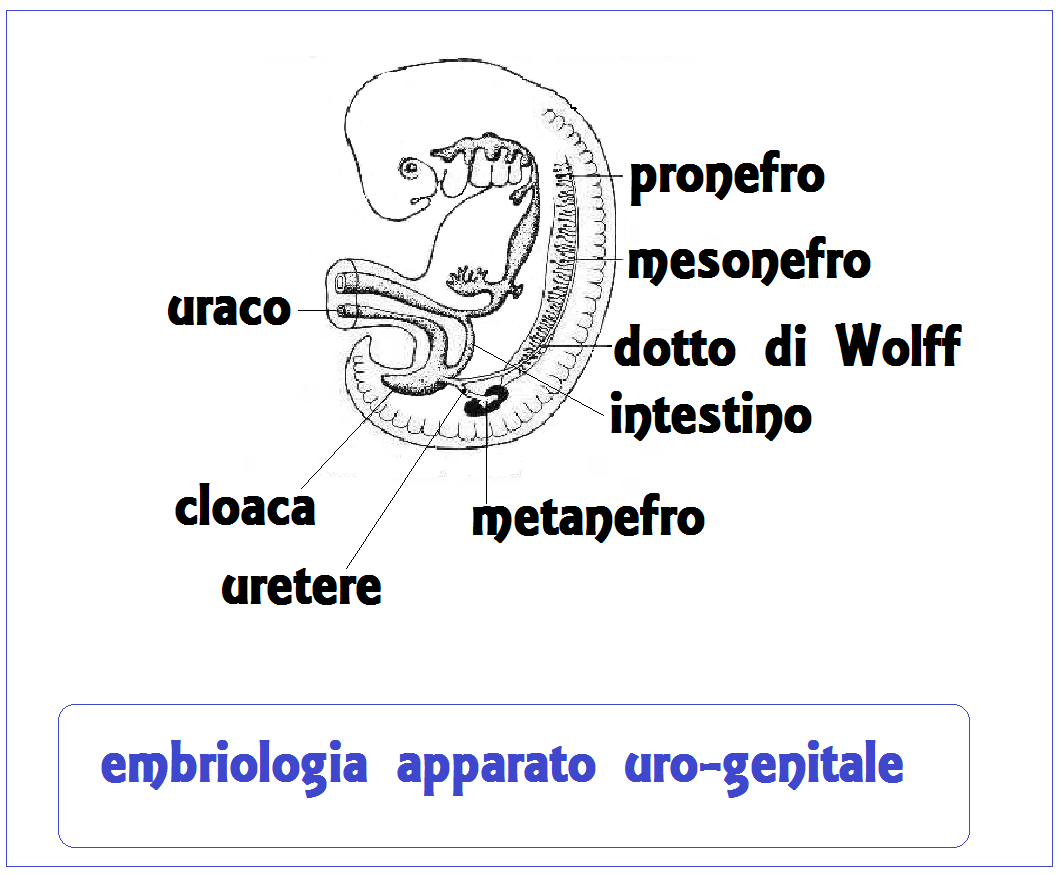
Il pronefro è costituito da circa 7 paia di canalicoli ricurvi, a convessità mediale, associati ai somiti dal 7° al 14°. L’estremità distale di questi canalicoli si unisce con quella dei canalicoli successivi e forma un dotto collettore, dotto di Wolff, che raggiunge la cloaca, dove sbocca dopo averne perforato la parete.
Il mesonefro, o corpo di Wolff, si estende dal 9° somite (corrispondente al 6° segmento cervicale definitivo) fino al 26° somite (corrispondente al 4° segmento lombare). Esso compare verso la 3a settimana. di sviluppo embrionale (embrioni di 2,5 mm), raggiunge il massimo sviluppo fra la 4a e la 9a settimana, poi inizia un processo di regressione che si conclude verso la 16a settimana.
Nel periodo del suo massimo sviluppo il mesonefro è costituito da circa 30 paia di tubuli ricurvi ad S, disposti trasversalmente; uno dei loro estremi si mette in relazione con un nodulo vascolare arterioso (glomerulo), l’altro si apre nel dotto di Wolff, denominato anche per tale motivo dotto del mesonefro.
A differenza dei tubuli del pronefro, quelli del mesonefro non si aprono nel celoma; inoltre nel mesonefro il glomerulo è intero. Man mano che i tubuli nel mesonefro si sviluppano, lo spazio a loro disposizione nella parete del corpo embrionale diviene insufficiente; si forma così una cresta che sporge nel celoma ai due lati del mesentere dorsale, detta cresta urogenitale. Quasi contemporaneamente alla sua formazione (cioè in embrioni di 5 settimane e di lunghezza di 6 mm), detta cresta si suddivide longitudinalmente in una porzione più laterale o cresta mesonefrica, ed in una più mediale o cresta genitale costituita da una zona midollare ed una corticale. Le cellule germinali primordiali (protogoni) migrano, già dal 14° giorno di fecondazione, dal mesoderma del sacco vitellino sulle creste genitali, penetrano in esse e formano i cordoni sessuali.
Nel maschio i cordoni sessuali si formano soprattutto dalla proliferazione della midollare, mentre nella femmina prolifera soprattutto la corticale. Se l’embrione è geneticamente maschile, per l’influenza esercitata dal gene SRY presente sul braccio corto del cromosoma Y, i cordoni sessuali continuano a proliferare, si approfondiscono nella midollare e danno luogo ai tubuli seminiferi (1-3). Se invece l’embrione è geneticamente femminile, i cordoni sessuali si suddividono in una serie di ammassi cellulari costituiti prevalentemente da cellule della granulosa che circondano le cellule germinali primordiali.
Sino a tutta l’8a settimana di vita embrionaria, i due sistemi dei dotti Wolffiani e mülleriani rimangono uguali sia nel sesso maschile che in quello femminile; solo dopo l’8a w, con l’inizio della differenziazione sessuale, uno dei due sistemi comincerà a subire fenomeni di involuzione.
Nella donna il mesonefro e il sistema dei dotti mesonefrici o dotti di Wolff subiscono un’involuzione quasi completa per cui nella donna adulta abitualmente si riscontrono solo alcuni residui di tale struttura; precisamente la porzione superiore del mesonefro dà origine all’epoophoron o corpo di Rosenmuller, la porzione intermedia dà origine al paraovario o dotto dell’epoophoron e nella sua porzione inferiore può dare origine a tratti più o meno lunghi del canale di Malpighi-Gartner.
Nel maschio, invece, l’involuzione delle strutture mesonefrotiche è meno completa; il dotto di Wolff e parte dei tubuli mesonefrici si trasformano nei condotti genitali definitivi precisamente dai tubuli mesonefrici derivano i vasi efferenti dell’epididimo e dal dotto do Wolff derivano il dotto dell’epididimo e il dotto deferente.
Il metanefro rappresenta l’abbozzo del rene definitivo. Esso deriva dalla porzione più caudale della corda nefrogena. Il sistema del canale di drenaggio (uretere, bacinetto renale, calici, dotti papillari e tubuli collettori e retti) deriva da un diverticolo della porzione terminale del dotto di Wolff; l’unità secretoria o nefrone deriva invece dai tessuti della corda nefrogena, in maniera simile a quella descritta per il mesonefro. Però contrariamente a quanto si verifica nel mesonefro, ogni tubulo secretorio metanefrico sbocca separatamente in un tubulo collettore del sistema di drenaggio sopra citato.
Dotti di Muller: i dotti genitali della femmina derivano da un abbozzo autonomo, il sistema dei dotti di Muller (dotti paramesonefrici), che compare verso la 6a settimana di sviluppo. I dotti di Muller si formano anche nel maschio ma vanno poi incontro a degenerazione verso la 12a sett. nella femmina invece evolvono progressivamente, dando origine alle salpingi, al fondo e corpo dell’utero, ed al 1/3 superiore della vagina. I dotti di Muller derivano da un solco che compare nell’embrione di 6 settimane (CRL 10 mm) sulla faccia laterale della porzione craniale di entrambe le creste urogenitali; il condotto vero e proprio si forma quasi subito: i bordi del solco iniziale procedono caudalmente, quindi si saldano trasformando il solco in un canale. Tale canale si prolunga poi in senso caudale per la progressione e la crescita della sua estremità a fondo cieco, decorrendo subito sotto l’epitelio celomatico e lateralmente al dotto di Wolff.
L’organogenesi vera e proprio dell’apparato genitale inizia quindi sul finire della sesta settimana dalla fecondazione (8a di amenorrea). La differenziazione della gonade primitiva in ovaio o testicolo è controllata geneticamente. Infatti è la presenza del cromosoma Y che determina il formarsi del testicolo, mentre la presenza di due cromosomi X determinano il formarsi delle ovaie. L’abbozzo del testicolo è già evidente alla settima-ottava settimana, mentre è solo tra la undicesima e la diciassettesima w che l’ovaio è ben delineato. Per la formazione del testicolo occorre, come si è detto, la presenza del cromosoma Y e i geni che determinano lo sviluppo di questo tipo di gonade sembra che siano localizzati nella regione centromerica, forse nel braccio corto del cromosoma Y.
Antigene HY: Tra i fattori che inducono la formazione del testicolo è stata data anche importanza ad una proteina denominata antigene HY che si formerebbe sulla superficie cellulare a causa della presenza del cromosoma Y. Quando questa proteina viene incorporata nelle cellule della gonade primitiva si formerebbe il testicolo; in assenza di questo antigene (e cioè in assenza del cromosoma Y), come nel caso di cariotipo femminile normale lo sviluppo della gonade indifferenziata evolve verso l’ovaio. La teoria dell’Histocompatibility-Y (antigene HY) è stata di recente criticata perchè non spiegherebbe come mai gli individui 45, X abbiano l’antigene in questione, per quanto a basso titolo. E’ stato perciò ipotizzato che la X contenga loci capaci di sopprimere l’HY oppure che il locus per l’HY non sia situato solo sulla Y, ma anche su un autosoma o anche sulla X.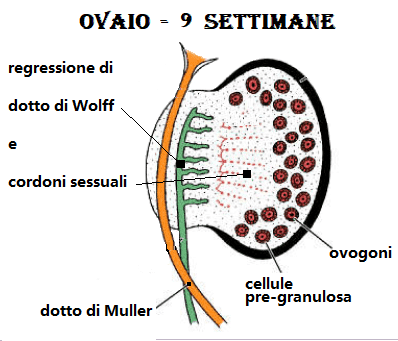
Si deve tener presente che per formare due ovaie normali occorrono due cromosomi X, pure normali. Se esiste un solo cromosoma X (45,X) si formano due ovaie, ma si assiste poi alla rapida distruzione (atresia) di tutti i follicoli per cui ne risultano due gonadi disgenetiche rappresentate per lo più da una  strisca di tessuto fibroso (“Streak gonad”).
strisca di tessuto fibroso (“Streak gonad”).
L’abbozzo gonadale indifferenziato, è rivestito esternamente da un epitelio cubico (il cosiddetto epitelio superficiale dell’ovaio) e costituito internamente da un tessuto lasso, detto blastema, che in gran parte deriva dalla proliferazione delle cellule dell’epitelio superficiale che si approfondano nel mesenchima sottostante. Solo nel corso della 7a sett. la gonade assume caratteri distintivi specifici nei due sessi.
Per quanto riguarda le cellule germinali esse verosimilmente derivano da alcuni elementi che si differenziano dalle cellule somatiche già prima che inizi la organogenesi dell’embrione. Queste cellule, dette cellule germinali primordiali, sono reperibili nel contesto dell’endoderma che riveste il sacco vitellino, caudalmente al disco embrionale. Le cellule germinali migrano in seguito verso l’embrione, lo raggiungono a livello della cloaca e proseguono la loro migrazione in senso caudo-craniale, lungo la parete dorsale dell’intestino primitivo ed il mesentere dorsale, fino a raggiungere la zona mesodermica dove si sta sviluppando la cresta genitale. Molti sostengono che le cellule germinali primordiali sono le uniche progenitrici di tutte le cellule germinali definitive; non si esclude invece da parte di altri la possibilità che un certo numero di cellule germinali definitive derivi localmente per differenziazione dell’epitelio superficiale.
Nelle femmine durante la 7a w, la gonade riduce la sua base d’impianto fino a formare l’abbozzo del mesovario, ma non presenta significative variazioni nella struttura del blastema; Nel corso dell’8a w di sviluppo il blastema indifferente della gonade femminile si differenzia in una zona compatta periferica o corticale primitiva, ed in una zona più lassa al centro, midollare primitiva. In ambedue le zone sono distinguibili numerosi piccoli ammassi di cellule blastematose (cordoni sessuali primari) che circondano una o più cellule germinali primitive dette, ora, ovogoni. Sempre a quest’epoca, infine, nella midollare primitiva si estende una propaggine di cellule derivate dai cordoni sessuali primari che invade parte del mesoovario e dà origine alla rete ovarii primitiva.
Nel corso del 3° e del 4° mese di sviluppo si verifica la ristrutturazione definitiva dell’ovaio. L’ovaio aumenta notevolmente di volume per la formazione della cosiddetta corticale definitiva (o corticale secondaria), che deriva dalla proliferazione di cellule della corticale primitiva il cui accentuato sviluppo caratterizza la formazione della gonade femminile, nonché dalla proliferazione di cellule dell’epitelio superficiale. Più tardi, verso il 6° mese, si formano dei sottili setti connettivali, a partenza dalla regione della rete ovarii, che suddividono la sostanza della corticale definitiva in tratti cordoniformi (cordoni o lobuli corticali). Questi setti connettivali, infine, si espandono alla periferia subito al di sotto dell’epitelio superficiale a formare uno strato sottile e continuo, detto tunica albuginea dell’ovaio.Contemporaneamente alla formazione della corticale secondaria si verifica la maturazione della midollare primitiva, che viene sostituita da uno stroma fibroso e vascolare che costituisce la midollare definitiva. Durante gli ultimi mesi della vita fetale nella corticale secondaria i lobuli cordoniformi già descritti si frammentano per la comparsa di numerosi setti connettivali che separano gruppetti di ovogoni e singoli ovociti gli uni dagli altri. Attorno a tali ovociti prima della 20a settimana si differenzia una capsula monostratificata di cellule epitelio simili e così si giunge alla formazione del follicolo primordiale.
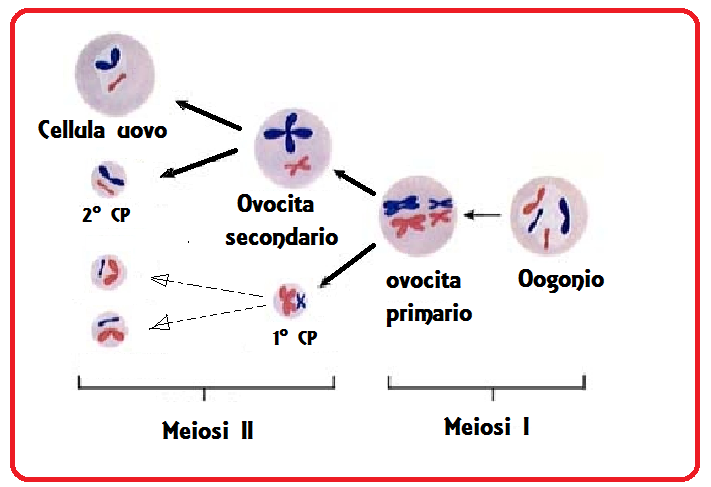
Meiosi/mitosi: Gli ovogoni sono le cellule con 46 cromosomi (diploidi, euploidi) che si moltiplicano dividendosi mitoticamente. Verso la undicesima settimana taluni ovogoni cominciano a trasformarsi in ovociti primitivi perché in essi inizia il processo di meiosi, ma altri ovogoni continuano a moltiplicarsi per mitosi. Si hanno, infatti, dapprima diverse divisioni di tipo mitotico, per mezzo delle quali il numero delle cellule germinali, valutato intorno a 700-1300 durante la migrazione, aumenta enormemente una volta raggiunta la struttura ovarica in via di formazione, fino a raggiungere il numero di circa 600.000 durante il secondo mese ed un massimo di circa 7 milioni verso il 5° mese di vita intrauterina. Dopo tale periodo il numero degli elementi germinali va incontro ad una notevole riduzione, sia per la cessazione delle mitosi negli ovogoni e sia per il determinarsi di processi di involuzione che colpiscono gli elementi germinali in vari stadi del loro sviluppo (atresia follicolare). Circa 5 milioni di cellule germinali sono eliminate così tra il 5° mese ed il termine della vita intrauterina e, dei 2 milioni di cellule restanti, oltre la metà mostra già segni di atresia. Alla nascita si contano circa 700.000-2.000.000 di follicoli primitivi. Al momento della pubertà gli ovociti rimasti sono 100.000-300.000.
Il processo meiotico si divide in due stadi: Meiosi 1 (M1) e Meiosi 2 (M2). Nello stadio M1 si verifica la separazione dei cromatidi. A seconda dell’aspetto che assumono i cromosomi, il processo meiotico viene suddiviso in quattro diverse fasi denominate profase, metafase, anafase e telofase. A sua volta la profase della prima divisione meiotica si compone di cinque periodi:
– leptotene: i cromosomi divengono visibili; precedentemente è avvenuta la replicazione del DNA, pertanto ogni cromosoma consiste di due cromatidi giustapposti;
– zigotene: i cromosomi sono chiaramente individuabili all’interno del nucleo; quelli omologhi si avvicinano e si appaiono progressivamente, con un movimento che ricorda quello di una cerniera lampo;
– pachitene: l’appaiamento dei cromosomi diviene ancora più intimo ed essi iniziano a scambiarsi reciprocamente alcuni tratti, mediante un fenomeno chiamato “crossing over”;
– diplotene: i cromosomi omologhi tendono a separarsi, restando uniti solo in alcuni punti, detti chiasmi, dove i cromatidi sono disposti in modo tale da formare una X, e che corrispondono ai tratti in cui avviene lo scambio di materiale genetico;
– diacinesi: i cromosomi appaiati tendono a migrare verso la periferia del nucleo.
Nel corso del periodo di diplotene il processo meiotico subisce nel feto femminile un arresto di durata molto lunga ed il passaggio al successivo periodo avviene dopo molti anni e cioè al momento della pubertà (4).
Nel maschio, invece, la meiosi inizia solo dopo la pubertà, e si verifica senza interruzione, in modo continuo.
Alla pubertà gli ovociti primitivi riprendono progressivamente e ciclicamente, in occasione del processo ovulatorio, l’evoluzione della Ia divisione meiotica interrotta allo stadio di diplotene della profase. Si riprende quindi con la metafase, in cui si osserva la rottura della membrana nucleare e la formazione del fuso, costituito da fasci di fibre proteiche orientate in senso longitudinale. I due centrioli vanno a collocarsi uno dalla parte di un polo e l’altro dalla parte del polo opposto della cellula, e su di essi convergono le fibre del fuso, che dal capo opposto si connettono con i centromeri dei cromosomi. A questo punto il centromero di ogni cromosoma si sdoppia ed ogni centriolo trascina verso il proprio polo il cromatide corrispondente (anafase). Nella telofase i cromatidi giunti ai poli cellulari ripassano dallo stato di addensamento a quello più diffuso. Alla fine della Ia meiosi avremo un ovocita maturo e un 1° globulo polare.
Dopo l’ovulazione, se l’ovocita è penetrato dallo spermatozoo, avviene la seconda divisione meiotica che è essenzialmente uguale ad una divisione mitotica, con la differenza che il numero dei  cromosomi è aploide. per cui si formano due gameti (ovocita + 2° corpuscolo polare) contenenti 23 cromosomi ciascuno ma con notevole differenza di grandezza perchè l’ovocita secondario riceve quasi tutto il citoplasma presente nell’ovocita primario. La divisione citoplasmatica, non cromosomica, è disuguale per cui da un lato resta l’ovocita secondario, dall’altro viene espulso il 2° corpuscolo polare. Appena il 2° globulo polare viene espulso la maturazione dell’ovocita è completa e può completarsi il processo di fecondazione e formazione dell zigote che apparirà dopo 12 ore. L’ovocita maturo ha un diametro di 80-100 micron, i globuli o corpuscoli polari hano un
cromosomi è aploide. per cui si formano due gameti (ovocita + 2° corpuscolo polare) contenenti 23 cromosomi ciascuno ma con notevole differenza di grandezza perchè l’ovocita secondario riceve quasi tutto il citoplasma presente nell’ovocita primario. La divisione citoplasmatica, non cromosomica, è disuguale per cui da un lato resta l’ovocita secondario, dall’altro viene espulso il 2° corpuscolo polare. Appena il 2° globulo polare viene espulso la maturazione dell’ovocita è completa e può completarsi il processo di fecondazione e formazione dell zigote che apparirà dopo 12 ore. L’ovocita maturo ha un diametro di 80-100 micron, i globuli o corpuscoli polari hano un diametro di 10 micron. I globuli polari non sembrano assumere alcun ruolo nella fisiologia dell’ovulazione e della fertilizzazione ma il 1° corpuscolo polare costituisce un’ottima fonte di informazioni su eventuali mutazioni cromosomiche e geniche presenti nel gamete femminile e che in parte ha sostituito la diagnosi pre-impianto dell’embrione (PGD). Infatti se la patologie. Gli oociti ottenuti dopo Pick-Up vengono posti sotto un microscopio con micromanipolatore e raggio laser freddo. Il fascio laser colpendo tangenzialmente l’oocita nella zona pellucida produce un’apertura, attraverso la quale un microago aspira il 1° corpuscolo polare e lo depone in una provetta a pareti sottili (Thin Wall) dove verrà analizzato nel laboratorio di Biologia Molecolare.
diametro di 10 micron. I globuli polari non sembrano assumere alcun ruolo nella fisiologia dell’ovulazione e della fertilizzazione ma il 1° corpuscolo polare costituisce un’ottima fonte di informazioni su eventuali mutazioni cromosomiche e geniche presenti nel gamete femminile e che in parte ha sostituito la diagnosi pre-impianto dell’embrione (PGD). Infatti se la patologie. Gli oociti ottenuti dopo Pick-Up vengono posti sotto un microscopio con micromanipolatore e raggio laser freddo. Il fascio laser colpendo tangenzialmente l’oocita nella zona pellucida produce un’apertura, attraverso la quale un microago aspira il 1° corpuscolo polare e lo depone in una provetta a pareti sottili (Thin Wall) dove verrà analizzato nel laboratorio di Biologia Molecolare.


la meiosi II, equazionale, post-fecondazione, divide il globulo polare primario in due globuli polari secondari e l’ovocita secondario in un ovotidio e un terzo globulo polare secondario. I globuli polari sono dei corpi degeneri, con poco citoplasma e pochi organuli. Vengono subito riassorbiti dalla mucosa.